

At the 2014 Bay Delta Science Conference, Matt Nobriga, a fish biologist with the U.S. Fish and Wildlife Service, gave this presentation on how fish utilize tidal marshes in the Delta. He began by noting that the despite a lot of research over the past decades, there is still much uncertainty regarding tidal marsh restoration.
The first issue of the San Francisco Estuary and Watershed Science was published in 2003; the subject was tidal wetlands. “And here we are 11 years later and there’s this concern that there’s still a lot of uncertainty,” he said. “And there always will be some; that’s life and the scientific endeavor. The response of California, as the 8th largest economy, was a very robust scientific response to that uncertainty as it stood at the time, and so in typical Bay-Delta fashion, although we have uncertainties, we probably have different uncertainties.
 But before he began his presentation, Mr. Nobriga had a comments to make about key uncertainties regarding tidal wetland/marsh restoration, acknowledging that these weren’t necessarily on topic with his talk. “The first one is that I think that tidal marshes and wetlands will produce fish and wildlife benefits,” said Mr. Nobriga. “They are a habitat feature of temperate zone estuaries all over the world. I think it will go without saying that something is going to benefit from large-scale marsh and wetland restoration if that is done in this system.”
But before he began his presentation, Mr. Nobriga had a comments to make about key uncertainties regarding tidal wetland/marsh restoration, acknowledging that these weren’t necessarily on topic with his talk. “The first one is that I think that tidal marshes and wetlands will produce fish and wildlife benefits,” said Mr. Nobriga. “They are a habitat feature of temperate zone estuaries all over the world. I think it will go without saying that something is going to benefit from large-scale marsh and wetland restoration if that is done in this system.”
“Secondly, bivalve grazing: it’s an eternal theme, and it’s one of the most excellently studied and personally interesting things to me,” he said. “But we know it’s a problem, so can we start talking about solutions to that problem? I would argue that we should try to think about that.”

“And then one last thing,” he said. “One of the key themes about my talk is we actually do know a lot of the fundamentals about fish and habitat in this system because we’ve been monitoring it and researching it rather extensively for decades. That has implications for the remnant tidal marshes and various microhabitats that exist out there right now, but if you were to do a lot of marsh restoration, or if you were to do a lot of wetland restoration, I don’t think anybody can really guess what emergent outcomes might come from that. So I just want to leave people with that thought. It’s not to say that nobody should think about it or try, it is just that I don’t know how well we could do that.”
Mr. Nobriga said in his presentation, he would make two main points:
- The fundamentals of fish habitat are known; more synthesis is needed: “I think that the fundamentals of fish habitat are known in this system, but that’s growing so fast – it’s about to quadruple in size in the next few years, making it all that much more harder to try keep on top of it. So there is a need for synthesis and there’s a need for fairly regular re-synthesis of that.”
- Freshwater flow regime is a potentially profound problem. “I’m going to play some games with freshwater flow regime because I think that this is something that conditions the fish assemblage that is there now and will continue to do so as we change the landscape and we need to think about it.”
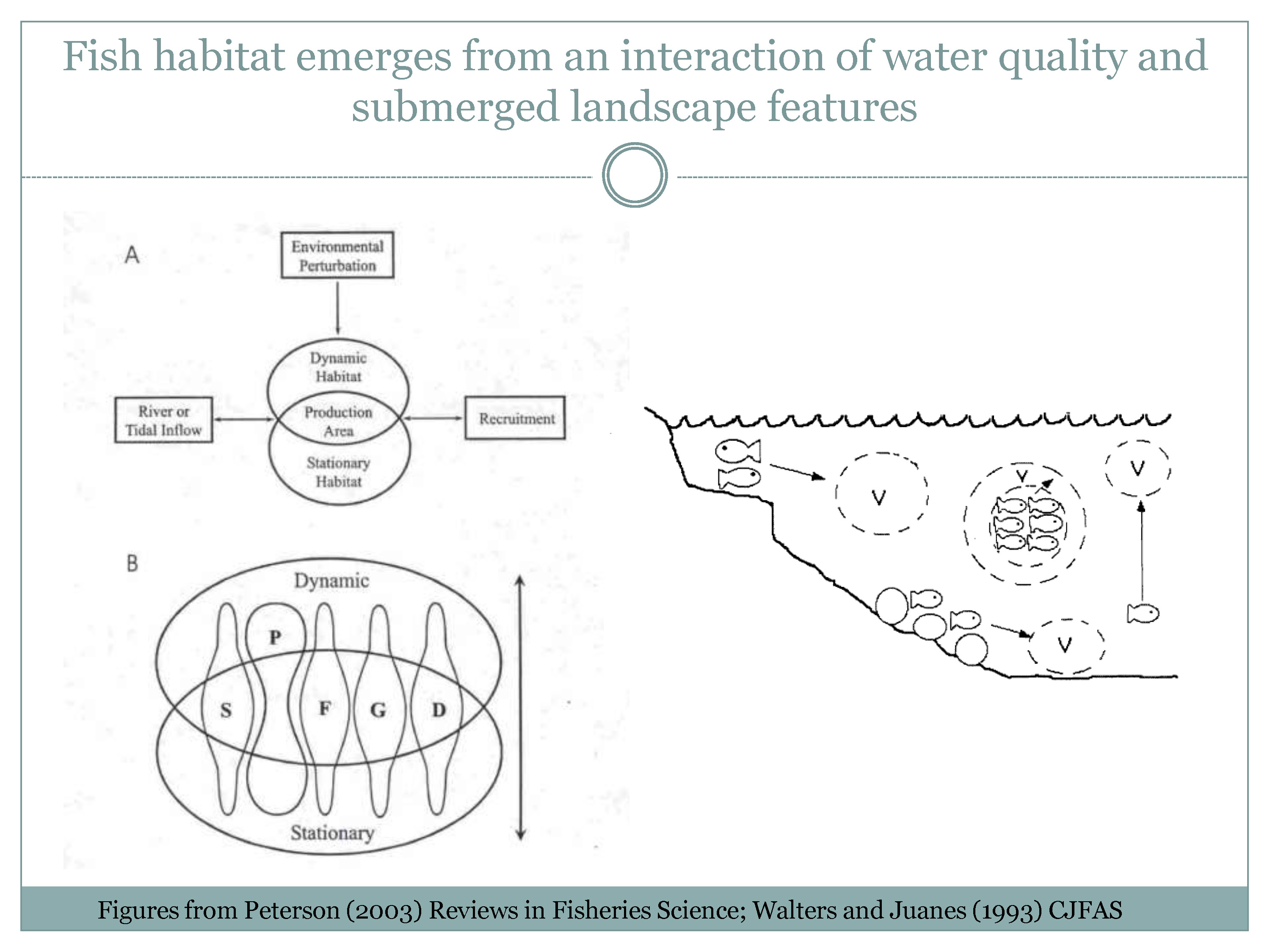 He then presented some basic fish conceptual models, noting that fish habitat emerges from an interaction of water quality and submerged landscape features. The first figure on the left hand side of the slide is from a paper by Mark Peterson in Reviews in Fisheries Science in 2003. “This one is actually about tidal river estuaries, which is what the northern San Francisco Estuary is,” said Mr. Nobriga. “So in panel A, there is a series of things that go on here – you have river tidal inflow and other environmental perturbations, and those two things create an intersection of your stationary habitat attributes, which is your landscape as it exists at the moment. They are not stationary forever, but they are stationary relative to dynamic habitat attributes which is the water quality of the estuary changing over that landscape. The intersection of that will produce production areas and those production areas will be different for different fishes and different life stages of even the same fish. And that will result in recruitment.”
He then presented some basic fish conceptual models, noting that fish habitat emerges from an interaction of water quality and submerged landscape features. The first figure on the left hand side of the slide is from a paper by Mark Peterson in Reviews in Fisheries Science in 2003. “This one is actually about tidal river estuaries, which is what the northern San Francisco Estuary is,” said Mr. Nobriga. “So in panel A, there is a series of things that go on here – you have river tidal inflow and other environmental perturbations, and those two things create an intersection of your stationary habitat attributes, which is your landscape as it exists at the moment. They are not stationary forever, but they are stationary relative to dynamic habitat attributes which is the water quality of the estuary changing over that landscape. The intersection of that will produce production areas and those production areas will be different for different fishes and different life stages of even the same fish. And that will result in recruitment.”
Panel B is showing the dynamic/stationary habitat interaction and recruitment, he said. “It says that where a fish’s or a fish life stage’s stationary and dynamic habitat attributes are properly overlapped, its foraging is going to increase, its growth is going to increase, its survival is going to increase, and all of that’s going to happen because predation rates on those fish are going to decrease,” he said. “Why the emphasis on predation? Fish die in droves. And most fish seem to get eaten most of the time unless they’re in really traumatic habitats. If you think about the fecundity of fishes, if a fish population was going to stay stable for a length of time, on average, 2 eggs out of however many it lays in its lifetime have to survive to keep that, to replace that fish, and most fish are laying thousands of eggs over their life cycle. So your odds aren’t good and most of those seem go away by predation.”
 A different version of it comes out of a paper by Carl Walters and Frances Juanes that’s about 20 years old. “Carl Walters attributed this to his at-the-time elementary school-aged son as a demonstration of just how elementary fish biology really is,” Mr. Nobriga said. “But what this is pointing out that helps make this point about foraging and predation more clear is that most fish don’t actually eat exactly where they hide. You have fish that hang out in schools. Some fish have to get to the edge of the school to eat; you’re not going to get any food in the middle of the school, so you’re taking a risk of predation by getting out on the edge. Some fish migrate vertically. If there’s a lot of food in the water, you can do that right before dawn and dusk or however you do it, and your risk to exposure to predators is pretty limited.”
A different version of it comes out of a paper by Carl Walters and Frances Juanes that’s about 20 years old. “Carl Walters attributed this to his at-the-time elementary school-aged son as a demonstration of just how elementary fish biology really is,” Mr. Nobriga said. “But what this is pointing out that helps make this point about foraging and predation more clear is that most fish don’t actually eat exactly where they hide. You have fish that hang out in schools. Some fish have to get to the edge of the school to eat; you’re not going to get any food in the middle of the school, so you’re taking a risk of predation by getting out on the edge. Some fish migrate vertically. If there’s a lot of food in the water, you can do that right before dawn and dusk or however you do it, and your risk to exposure to predators is pretty limited.”
“If there’s not a lot of food, you’ve got to go earlier and you have to take a bigger risk,” he continued. “The bigger risk means more cumulative predation. If you live in the weeds or the rocks and you have to go out, same thing; if you live in the shallows and you’ve got to go deep, same thing; if you’re a splittail and you’re hanging out deep and you want to go shallow, same thing. There’s a risk/reward.”
“We get in a lot of arguments in my world about density dependence and fish and how can a rare fish have density dependence,” he said. “This paper is great, because it’s a mathematical demonstration that you get density dependent-looking relationships from an individual fish’s having to balance its rewards, its getting enough energy, to reproduce in its life, versus a risk of predation. So it’s not algae limiting out their nutrients in a flask, or bacteria in agar culture. This is an individual-based thing that will look like density dependence at a population scale but actually is just individuals having to balance the need to get food with the need to not get eaten.”
 He then presented a slide of the Peterson model of the San Francisco Estuary, which plots salinity against population. He said that salinity is the most dynamic habitat attribute of any estuary, and it does affect the population. “Salinity affects a fish’s physiology,” he said. “If its physiology is affected, that’s going to affect its health, it’s going to affect how it behaves, and as a consequence it’s going to affect its population dynamics.”
He then presented a slide of the Peterson model of the San Francisco Estuary, which plots salinity against population. He said that salinity is the most dynamic habitat attribute of any estuary, and it does affect the population. “Salinity affects a fish’s physiology,” he said. “If its physiology is affected, that’s going to affect its health, it’s going to affect how it behaves, and as a consequence it’s going to affect its population dynamics.”
“This is the cumulative salinity distributions of fishes in trawl data,” he said, noting that in the areas where the salinity is higher, they tend to catch a lot of marine fishes, and on the freshwater side of the estuary, they catch more salmon, smelt, shad and others. He also noted that a few fishes, like longfin smelt, starry flounder and Pacific staghorn sculpin do have a more even distribution across the salinity gradient. “That’s a partitioning mechanism and that’s going to affect where in the estuary and where on that landscape this dynamic habitat is right for you.”
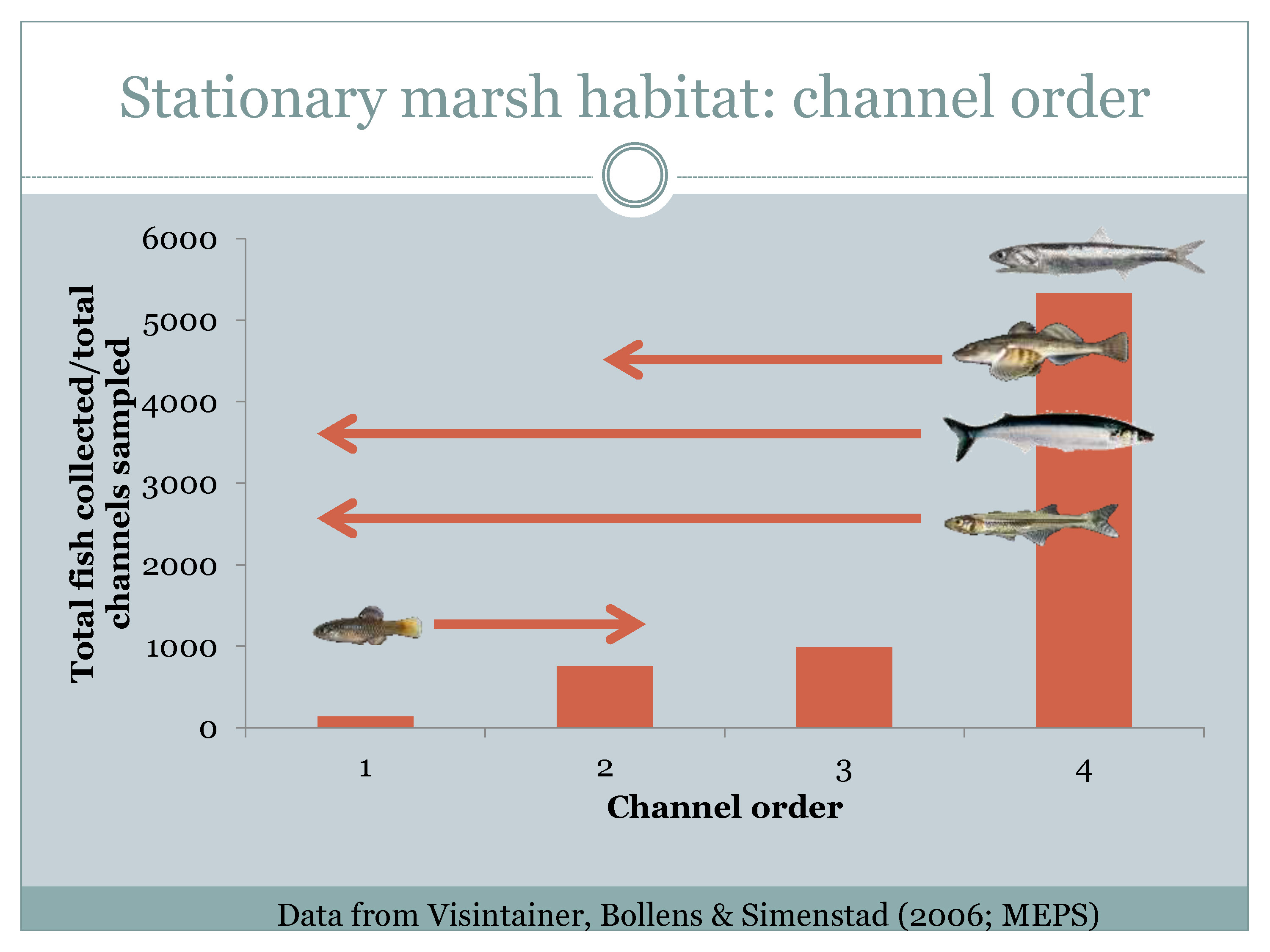 For the higher salinity side of the estuary, Mr. Nobriga next presented a graph that he redrew from a paper by Tammy Visintainer et al with channel order on the x-axis and number of fishes sampled on the y-axis. He noted that this study was done in the China Camp Marsh adjacent to San Pablo Bay, a small marsh with tiny channels up to 4th order streams. “What you see is that there’s more fishes as you get out toward the Bay,” he said. “This is a fractal thing, so a big marsh like Suisun Marsh will go to much higher channel order than 4. If you look at SFEI’s historical Delta, the Sacramento River or the San Joaquin River was a tidal marsh channel at a historical scale. And so we could probably assume that that would keep going.”
For the higher salinity side of the estuary, Mr. Nobriga next presented a graph that he redrew from a paper by Tammy Visintainer et al with channel order on the x-axis and number of fishes sampled on the y-axis. He noted that this study was done in the China Camp Marsh adjacent to San Pablo Bay, a small marsh with tiny channels up to 4th order streams. “What you see is that there’s more fishes as you get out toward the Bay,” he said. “This is a fractal thing, so a big marsh like Suisun Marsh will go to much higher channel order than 4. If you look at SFEI’s historical Delta, the Sacramento River or the San Joaquin River was a tidal marsh channel at a historical scale. And so we could probably assume that that would keep going.”
“So for restoration, from a fisheries perspective, what you’re trying to do is get productivity to the edge of the marsh, whatever the edge of your marsh is at the scale you’re talking about,” he said. He noted that in this marsh, there is some partitioning with the only species using the back being the non-native rainwater killfish. “On the other end, northern anchovy would use the 4th order channels but apparently didn’t penetrate the marsh any farther, reemphasizing that point as a pelagic planktavore that if you want to get at some food, you’ve got to get productivity to the edge of the marsh. We can haggle about whether we’re restoring marshes—you’ve got to be pretty specific if you’re restoring the interior of the marsh for a particular fish as opposed to transferring productivity to a place that most of the fishes that we think about are actually going to get to it.”
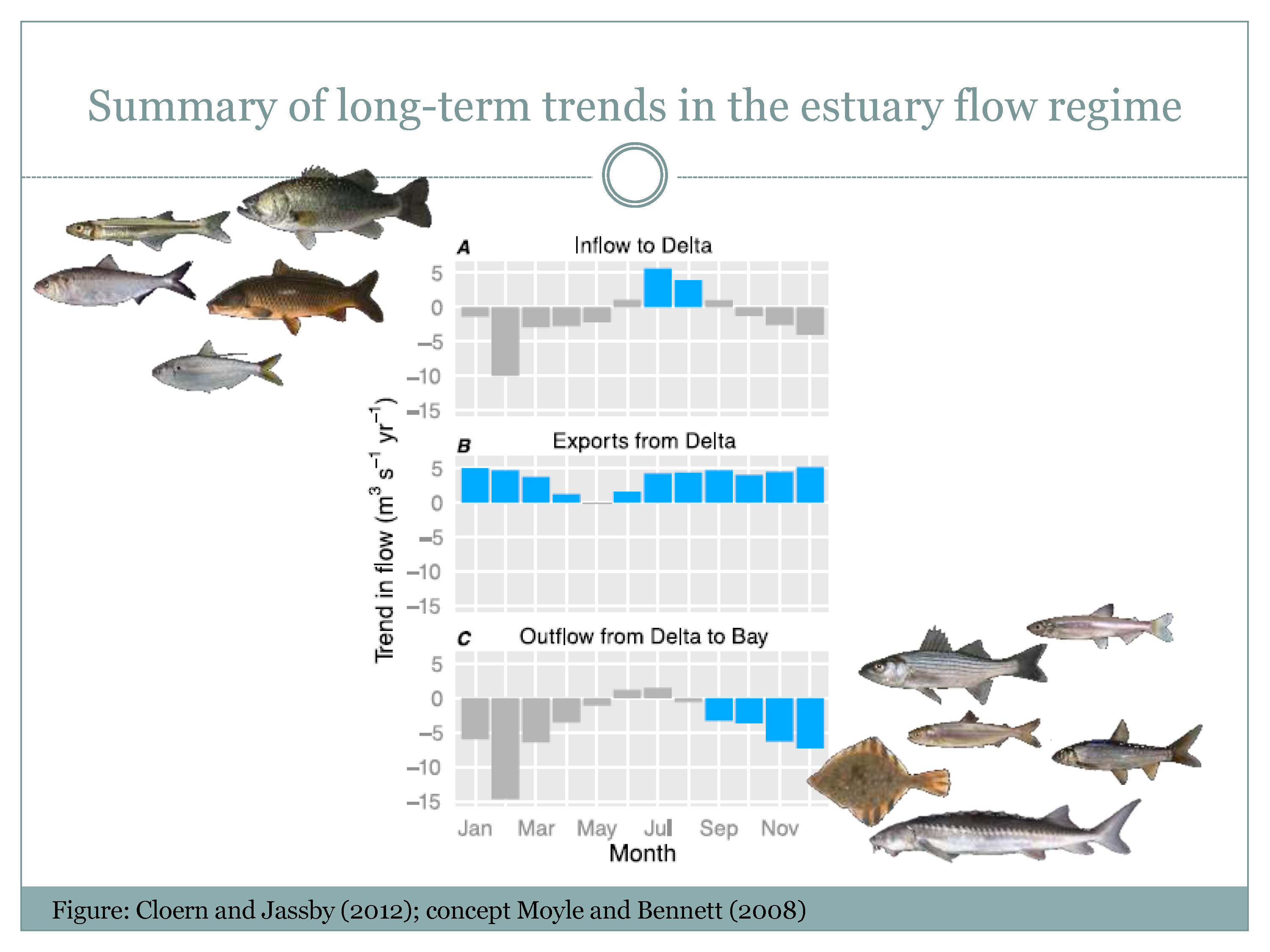 Mr. Nobriga then said he was going to do a thought experiment with the concept of a freshwater flow regime, presenting a slide of a summary of the long-term trends in the Delta’s flow regime from a recent paper by Jim Cloern and Alan Jassby. He noted that the x-axis is the have month of the year and the y-axis is the slope over time in the trend in these various hydrologic variables. “The top panel is Delta inflow; that has generally trended down most months of the year except in the summer, June through September,” he said. “Exports have generally increased—this is all since 1956—most months of the year except May, and Delta outflow has tended to trend down almost every month except a little bit of an uptrend perhaps in the middle of the summer. If it’s blue it’s a statistically significant trend and if it’s grey it’s not a statistically significant trend.”
Mr. Nobriga then said he was going to do a thought experiment with the concept of a freshwater flow regime, presenting a slide of a summary of the long-term trends in the Delta’s flow regime from a recent paper by Jim Cloern and Alan Jassby. He noted that the x-axis is the have month of the year and the y-axis is the slope over time in the trend in these various hydrologic variables. “The top panel is Delta inflow; that has generally trended down most months of the year except in the summer, June through September,” he said. “Exports have generally increased—this is all since 1956—most months of the year except May, and Delta outflow has tended to trend down almost every month except a little bit of an uptrend perhaps in the middle of the summer. If it’s blue it’s a statistically significant trend and if it’s grey it’s not a statistically significant trend.”
“If we think about this flow regime that we’re putting over the estuary’s landscape, we’re increasing mid-summer flows into the Delta, and we’re decreasing flows most of the year out to the estuary, and so the crux of this thought experiment is that we’re incentivizing the ecosystem for lowland freshwater fishes and disincentivizing it for estuarine fishes. This is a concept that actually is in the 2nd PPIC report technical appendix initially by Peter Moyle and Bill Bennett.”
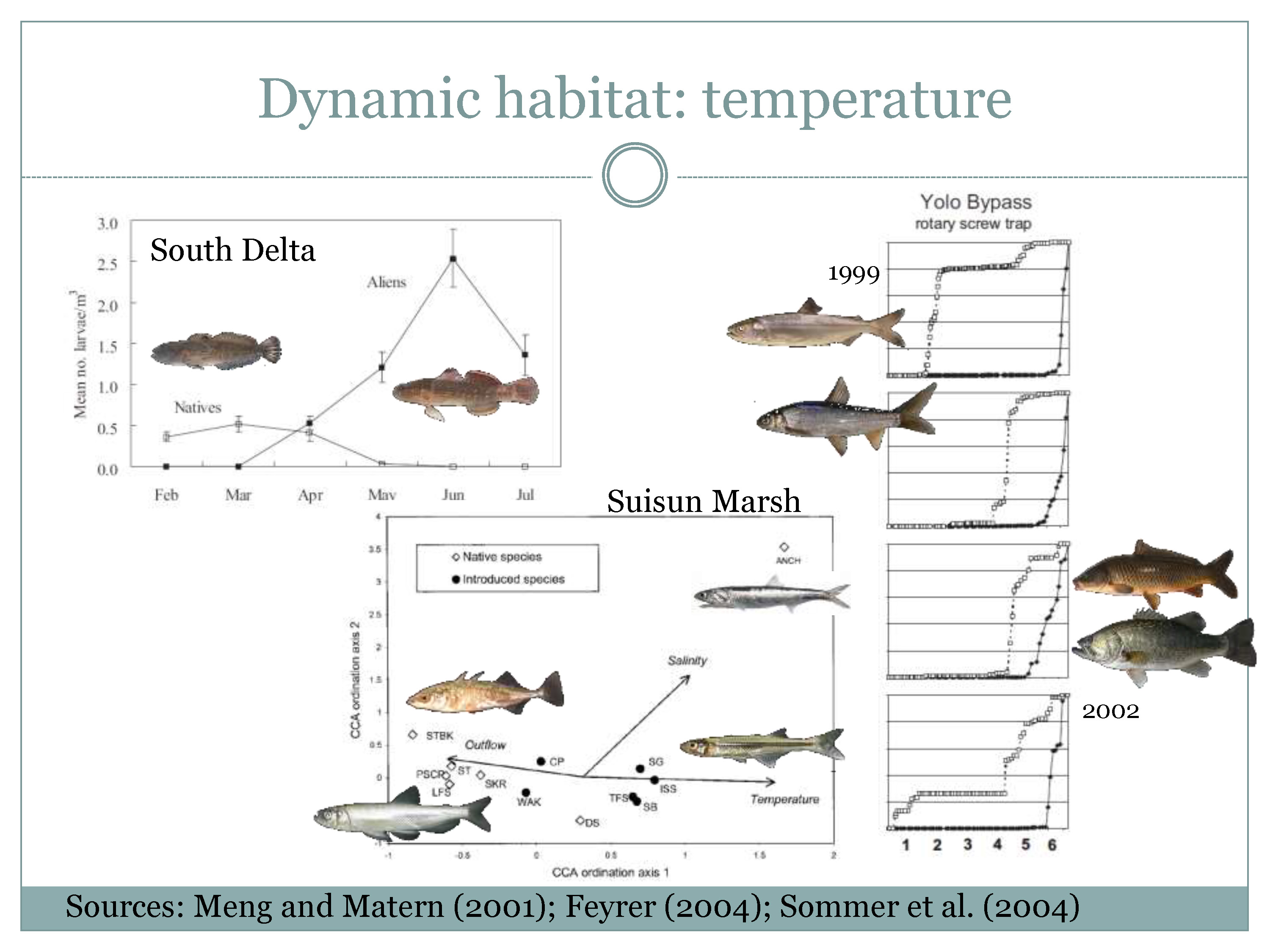 “So dynamic habitat in this part of the estuary is fresh or fresh-ish, and water temperature is pretty key to this conceptual model because most of the remnant native fishes spawn at colder water when the historical flow peaks occurred,” he said, noting that this is true for the south Delta, the Suisun Marsh and the Yolo Bypass.
“So dynamic habitat in this part of the estuary is fresh or fresh-ish, and water temperature is pretty key to this conceptual model because most of the remnant native fishes spawn at colder water when the historical flow peaks occurred,” he said, noting that this is true for the south Delta, the Suisun Marsh and the Yolo Bypass.
“I have to really tell you about my epiphany moment which was fishing the rotary screw trap in the Yolo Bypass in 1998,” Mr. Nobriga said. “It was a super wet year and it did something it doesn’t usually do – it was still flooding like crazy in June, April and May, and the screw trap’s catching salmon and then splittail and then all of a sudden in June as the temperature gets warmer, the same floodplain doing its thing is full of carp and largemouth bass. They require water too.”
 “That’s central to this conceptual model,” he said. “I built a generic fish out of a Ricker model. So, for people who aren’t familiar with that, it’s recruits, number at the next generation as a function of spawners, and some density dependence if you want it. So, I made both the recruits per spawner—how successful you were—a function of flow. More recruits when it’s wetter, less recruits when it’s drier; I increased the density dependence when it’s drier and let it ease up when it’s wetter because there’s more space. Sort of with my ‘aha’ moment in the Yolo Bypass.”
“That’s central to this conceptual model,” he said. “I built a generic fish out of a Ricker model. So, for people who aren’t familiar with that, it’s recruits, number at the next generation as a function of spawners, and some density dependence if you want it. So, I made both the recruits per spawner—how successful you were—a function of flow. More recruits when it’s wetter, less recruits when it’s drier; I increased the density dependence when it’s drier and let it ease up when it’s wetter because there’s more space. Sort of with my ‘aha’ moment in the Yolo Bypass.”
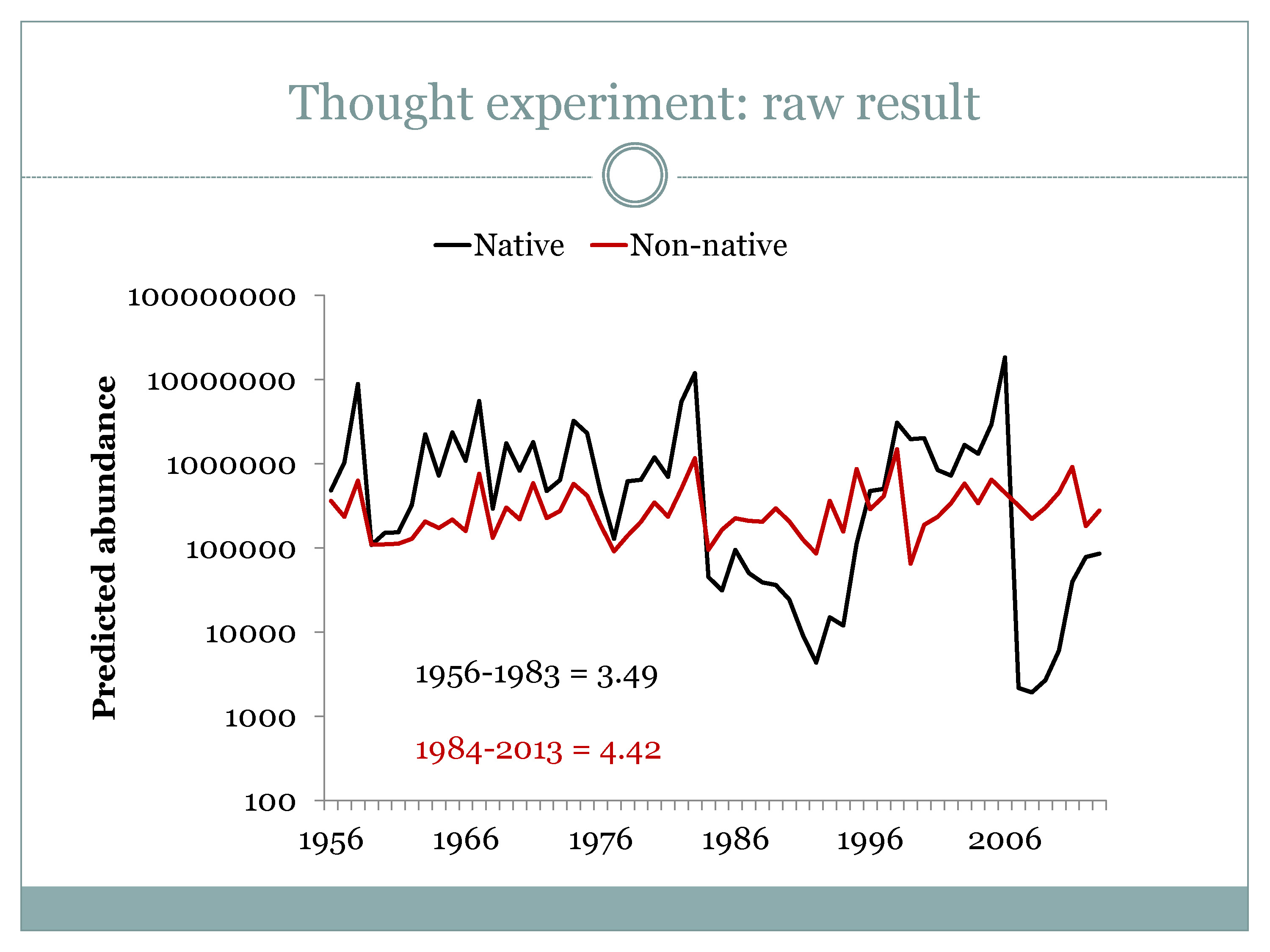 “The application here is I just applied this same exact mathematical fish to the historical flow time series,” he said. “The bottom line here is that for the first half of this time series, from the mid-‘50s to the mid-‘80s, the flow regime in this Ricker model predicts that the native fish would be 3½ times more abundant than the non-native fish at the median; and that since that time, that has actually reversed so that the non-native fish would be predicted to be 4½ times as abundant as the native fish. And the non-native fish doesn’t really have any trend; it just doesn’t do anything, whereas apparently the changes in flow are enough to influence the recruitment of a hypothetical native fish.”
“The application here is I just applied this same exact mathematical fish to the historical flow time series,” he said. “The bottom line here is that for the first half of this time series, from the mid-‘50s to the mid-‘80s, the flow regime in this Ricker model predicts that the native fish would be 3½ times more abundant than the non-native fish at the median; and that since that time, that has actually reversed so that the non-native fish would be predicted to be 4½ times as abundant as the native fish. And the non-native fish doesn’t really have any trend; it just doesn’t do anything, whereas apparently the changes in flow are enough to influence the recruitment of a hypothetical native fish.”
So what can we do? What should we do? “First, synthesize fish habitat information so it’s easier for a broad, multidisciplinary audience to digest,” he said. “Secondly, we need some broader thinking into this fish-flow relationship concept that goes beyond a spring X2 argument or a fall X2 argument. I think we need to have a suite of species that respond differently and that we know are physiologically different, and that we can understand what happens to their vital rates at different temperatures, different flows, different places, so that you can really begin to mechanistically understand how our choices as a society affect what we get as a fisheries ecosystem.”
“And also research clam control if you didn’t see the footnote,” Mr. Nobriga added.
- For the full power point: NOBRIGA_311Thur_PM2_BDSC2014
More from the 2014 Bay Delta Science Conference …
 Help Maven fill the funding gap and keep unique content like this flowing …
Help Maven fill the funding gap and keep unique content like this flowing …

Make a tax-deductible donation or “join the club” today!
